Why science favours lipless theropods
Despite what a plethora of enthusiasts claim; that ‘using Carr et al 2017 as a means to debunk said lips is flawed and shouldn’t be credible, as ‘crocodilians do not exhibit epidermal scales and instead keratinised skin.’ Is flawed.
This paper was unfortunately heavily misunderstood as Carr never said anything about ‘the same type of tissue being present’ as the paper was not about crocs but the analog was based on the presence of tactical sensors, as underlying for some sort of flat scales in tyrannosaurs or "mask".
So when Carr said: ‘As in crocs’ he and his team only acknowledge the presence of numerous of foramina on the premaxilla, maxilla and dentary and on the nasal as they then used as reference point ‘croc like face’.
But people got hung up and misunderstood this, so they wrote everywhere that Carr ‘claimed that crocs inhabit scales in their face and then Carr et al applied it as an analog for Tyrannosaurs’ which would be incorrect because ‘crocs don’t have scales’.
What many fail to realise is that Carr and his team DID compare tyrannosaur facial textures to ‘all’ archosaurs, consisting of 1 crocodilian species and 5 bird species.
‘In D.horneri and in ALL derived tyrannosauroids, the subcutaneous texture is coarse and shows the hierarchy of textures (Supplementary Discussion S3). In order to identify the soft tissue that produced this complex surface, we compare the condition of tyrannosaurids with that of crocodylians (Alligator mississippiensis) and birds (Struthio camelus, Anser sp., Anas sp., Cygnus sp., Meleagris gallopavo), and we followed several studies [29, 31, 39, 40] to identify osteological correlates imprinted on the cortical surface of facial bones to deduce their casual soft tissues.’
The study also mentioned that they used some other papers to help ‘create’ the facial integument and croc-like appearance.
(-Hieronymus, Witmer, Tanke, Currie 2009: The facial integument of centrosaurine ceratopsids: morphological and histological correlates of novel skin structures.
-Hieronymus, Witmer 2010: Homology and evolution of avian compound rhamphothecae.
-Papp 2000: Critical Appraisal of Buccal Soft-tissue Anatomy of Ornithischian Dinosaurs.
-Sedlmayr 2002: Anatomy, Evolution, and Functional Significance of Cephalic Vasculature in Archosauria.)
Carr et al - as late as 2017 with their team worked closely with Hieronymus, T. L. & Witmer, L. M. Homology to establish the various textures (osteological) The oft-cited and "maligned" Carr et al. 2017 study provides another essential premise.
The dentary exhibits the type of texturing that corresponds with armor-like keratinous integument seen in crocodiles, ceratopsians, and also on the dorsal part of the snout of tyrannosaurids. Whatever soft tissue grew there was fused closely to the bone and this is what makes up their epidermal tissue along and all the way to the alveloar process, not only on the maxilla, but the dentary top and bottom.
There was no other type of texture that would indicate some different type of facial tissue, as nothing in the bones indicated such presence. (Which is why squamata corresponding with extra oral tissue was excluded from the study). From this paper we can set the null hypothesis.
Regardless of whether you agree with the results of that study in terms of its final reconstruction, it established and collates a few crucial facts about tyrannosaurid facial integument.
First, textured bone on the faces of crocodylians, tyrannosaurids, ceratopsians, and birds alike corresponds with close proximity of integument to the bone. Secondly, the presence of ISOs is indicated to be common throughout tyrannosaurids.
It means this model actually addresses the relationship dinosaurs have to birds as well as it stated tyrannosaurs have certain structures present on their face that would go on to be more evident in bird species in other earlier dinosaurian lineages, hence the favour of lipless theropods doesn’t rely on just one study.
This is also not the only study discussing the presence of integumentary sensory organs in theropods.
Barker et al. 2017 studies the foramina of Neovenator and again compared it to a crocodiles as well as Spinosaurus and Baryonyx.
The study suggests the foramina in neovenator representing abudance of trigeminal nerves would be similar to those found in extant crocodilians (by ct scan). The conclusion suggested both the foramina and the internal nerve system would in fact correlate to a tactile hyper sensitivity akin to crocs.
Most significantly, the lower jaw shows evidence for neurovasculature that is also seen in birds.
Fig. 3
Complex neurovasculsture shown surrounding infilled dental alveoli of the maxilla of neovenator
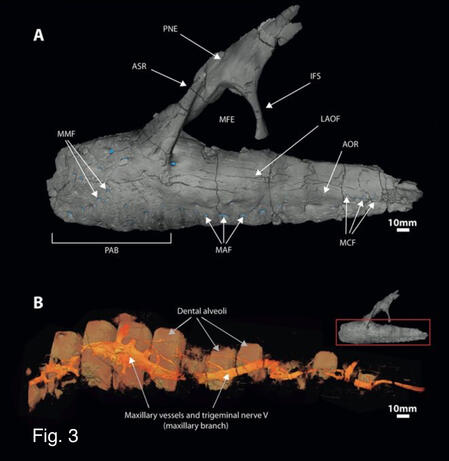
This crocodile like adaptation in daspletosaurus and neovenator is believed to have been applied in nesting behaviour, communication, defleshing etc.
(Ibrahim et al.2014 also once again establishes the commonality of ISOs in theropods. From a ct scan, s.aegyptiacus bears a similar amount of neurovascular foramina that is alike to a crocodile.)
Despite what multiple ct scans have established, some may counter argue that such hyper sensitivity would have been a burden as crocodilians use this trait underwater, unlike theropods. This of course is flawed as the opposition claimed that the ISO’s aid in detection of air movement, something that hasn’t been specified in any study. (The adaptation is more to be used in communication and hunting other than detecting air movements).
A recent study on Juravenator also shows epidermal scales located on the tail housing ISOs.
‘Although epidermal scales have been noted previously
on the tail of Juravenator, we report a unique scale type with distinctive circular nodes that we identify as integumentary sense organs, analogous to those in modern crocodylians. The surprising presence of such structures suggests the tail had a sensory function, which is nevertheless congruent with the inferred ecology of Juravenator and the evolution of integumentary sense organs among archosaurs.’
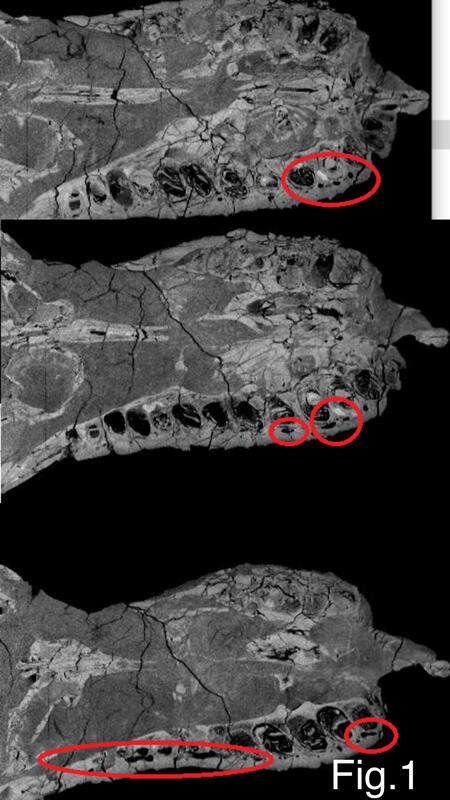
From this we can again conclude that ISOs are a common trait along theropods ranging from tyrannosaurids to carnosaurs and spinosaurs. (Fig 1 shows highlighted areas showing fairly big channels akin to what was reconstructed for Neovenator within Sue's left maxilla.)
A while ago there was a study based on the tooth structure in crocodilians and how it corresponds with their bone-crushing bite and tooth replacement cycle. What this has concluded is that crocodilians have very thin enamel aiding in their bite force and tooth replacement. They concluded that animals with this kind of tooth structure tend to keep enamel levels low as to benefit their tooth replacement cycle. Crocodilians tend to replace their teeth several times a month which is alike to theropods, which do so in and around the same time period.
What the study also did was scan crocodile and other archosaur teeth including tyrannosaurus teeth in order to compare and contrast the structure.
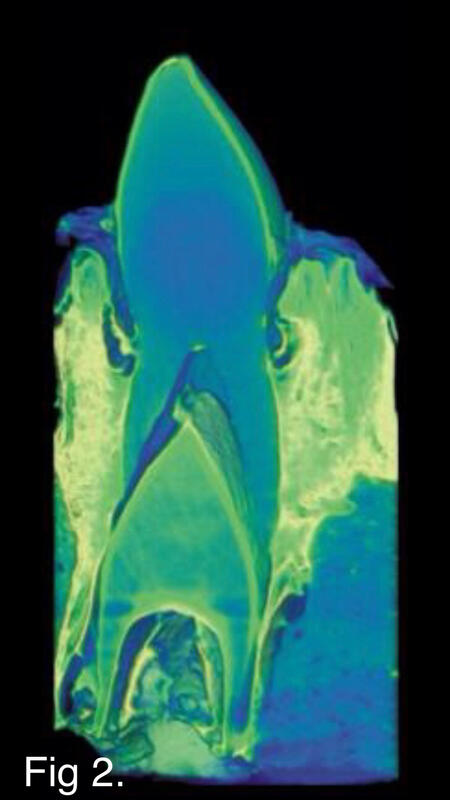
Fig. 2
‘The team used a three-dimensional x-ray scanner to measure the thickness of tooth enamel in crocodiles. They found regardless of tooth position -- incisor, canine, molar -- age or diet, crocodiles do not have thick tooth enamel. With this new information, the team also studied published data on dinosaur teeth and found that the data nearly matched what they were seeing in crocodiles. ‘It is particularly noteworthy that enamel in two tyrannosaurid teeth was nearly the same thickness as the enamel of a much smaller Allognathosuchus tooth and several teeth from Alligator. The apparently-thicker enamel in crocodyliforms may be related to the characteristic hard-biting feeding behavior.’
What this ultimately tells us is that once again, just like with facial integument, theropods exhibit extreme similarities with crocodilians.
Now that we know that theropods and crocodiles have the ‘same’ tooth structure, it would be only logical that their common ancestors evolved this trait possibly to aid in a ‘bone crushing’ bite force. In fact, the researchers at the University of Missouri have found that even their plant-eating ancestors had thin tooth enamel.
Now one argument that floats around is that tooth enamel requires moisture to be maintained. In actual fact, animals with exposed teeth such as elephants with their tusks, do have thin enamel on the tusks, likewise with musk deer and wild boars.
They are left exposed to the elements and this is probably the reason why the amount of enamel is kept low.
Crocodiles may be primarily aquatic but some species (like crocodylus palustris) estivate, directly avoiding the need to hydrate the enamel in contrast to the claim that they can only remain lipless due to the fact they spend so much time in water. Freshwater crocodiles do not appear to have any specific adaptations for estivation, but given an adequate refuge, they can survive many months without access to water as well.
The reason for this adaptation in crocodiles and theropods is because of the constant tooth shedding that would burden the need for thicker enamel. Enamel takes time to form, something that animals wouldn’t do constantly given the fact that these tooth replacement cycles last a month or so, hence being inefficient.
Theropods like crocodiles also tend to lead brutal lives, their teeth go through sheer amounts of abuse it’s no wonder the tooth replacement cycle is relatively fast. Many fossils show face biting to be a common behaviour among this group of dinosaurs.
In conclusion both theropods and crocodiles have the same thickness of enamel and both constantly replace their teeth through the course of their lives. Crocodiles do not require moisture for their enamel and the same can be assumed for theropods.
-Hieronymus, Witmer, Tanke, Currie 2009: The facial integument of centrosaurine ceratopsids: morphological and histological correlates of novel skin structures.
-Hieronymus, Witmer 2010: Homology and evolution of avian compound rhamphothecae.
-Papp 2000: Critical Appraisal of Buccal Soft-tissue Anatomy of Ornithischian Dinosaurs.
-Sedlmayr 2002: Anatomy, Evolution, and Functional Significance of Cephalic Vasculature in Archosauria.
-Witmer, L.M. and Ridgely, R.C. (2008), The Paranasal Air Sinuses of Predatory and Armored Dinosaurs (Archosauria: Theropoda and Ankylosauria) and Their Contribution to Cephalic Structure
-Christian, Keith, et al. “Some Physiological Consequences of Estivation by Freshwater Crocodiles, Crocodylus Johnstoni.” Journal of Herpetology
-Carr, T., Varricchio, D., Sedlmayr, J. et al. A new tyrannosaur with evidence for anagenesis and crocodile-like facial sensory system
-Barker, C.T., Naish, D., Newham, E. et al. Complex neuroanatomy in the rostrum of the Isle of Wight theropod Neovenator salerii .
-Ibrahim, Nizar & Sereno, Paul & Dal Sasso, Cristiano & Maganuco, Simone & Fabbri, Matteo & Martill, David & Zouhri, Samir & Myhrvold, Nathan & Iurino, Dawid. (2014). Semiaquatic adaptations in a giant predatory dinosaur.
“Crocodile-like sensory scales in a Late Jurassic theropod dinosaur” by Phil R. Bell and Christophe Hendrickx
-K.C. Sellers et al. The significance of enamel thickness in the teeth of Alligator mississippiensis and its diversity among crocodyliforms. Journal of Zoology
-Gignac, P.M., Erickson, G.M. The Biomechanics Behind Extreme Osteophagy in Tyrannosaurus rex
No matter how closely related and similar crocodilians and dinosaurs are, many modern paleoart depicts theropods possessing lizard-like traits. This isn't inherently wrong but from a scientific standpoint, the lizard-like depictions don't hold up.
Theropods and squamata are 2 distinct groups with varied differences between them. They have such different lifestyles and behaviours apart from physically contrasting traits, it's no wonder why the slow, dumb, cold-blooded, lizard-like depictions were abandoned after dinosaurs started becoming more understood.
It seems like these lizard qualities have resurfaced in paleoart today.
Anatomical differences between lizards and theropods are as follows;
Squamates possess labial glands that are covered by extra-oral tissue. They have relatively large circular foramen arranged in neat rows across the dentary and maxilla, occluding teeth that are small, thin and almost always covered by extra-oral tissue and of course, are extremely far from theropods phylogenetically. As well as that, there is no real consensus as to why lizards have extra-oral tissue.
Not to mention, theropods and squamates have an entirely different skull shape and anatomy, the differences which I will get to in a bit.
Now onto mammals.
Mammals are the only animals that have lips. Other animals have extra-oral tissues of different designs. It's important not to fall into the habit of generalization or colloquial speech when delving into the details. Mammals, have lips for different reasons, communication, food manipulation and suckling are just some of the uses for them.
To get the point across, lips are rendered useless in theropods. They obviously don’t need them in terms of communication, suckling etc etc, and don’t have labial glands.
Many archosaurs have beaks or beaklike structurs, both within and outside of dinosaurs. For instance, silesaurus, effigia, aetosaurs, pterosaurs, and some other archosaur taxa had beaks or beaklike structures.
Theropods repeatedly evolved beaks (oviraptorids, ornithomimids, limusaurus, therizinosaurs, and probably several times in birdline maniraptorans), for instance, which would suggest they all ancestrally had the genes to easily evolve true beaks.
Considering there are no beaked mammals (the bills of monotremes are not beaks), snakes, or lizards, this would suggest that having lips is a barrier to evolving beaks. (I will also point out that the foramina and skull texture in certain birds,
Insert pic
is similar enough to that seen in many toothed theropod clades to indicate that a keratinous covering is more plausible than fleshly lips.
On the topic anatomy, yes, crocodiles have a different tooth row compared to theropods. But do any lipped mammals have such extreme overbites as the likes of theropods? Many lipped reptiles have a ‘jaw gap’ for their teeth to fit. This prevents them from fully shutting their jaws and instead, extra-oral tissue account for this. In the case of some lipless reptiles their teeth will fit relatively smoothly between the maxillae as they lack a jaw gap. Some even have palet gooves to protect the lower teeth.
I might also point out, regarding tooth size, that the margins of the maxilla and dentary in croc monitors are concave, so as to accommodate larger tooth sizes behind lips. In theropods, we get the opposite extreme, and that convexity is important for tyrannosaurids' habit of eating bones.
Many theropods tend to grow to gargantuan sizes, and with more mass the problem with overheating arises. Researchers at Witmer lab have established the certitude that theropods such as majungasaurus crenatissimus had antorbital air sinuses that acted as a mechanism that cooled blood going into the brain. Large theropods affected by this thermal problem would flex their jaws to force air in and out of the sinuses causing moisture to evaporate and heat to be vented. This would have happened very frequently almost constantly with the possible exception of inactivity.
In this case, large theropods did have to keep their mouths ventilated, especially during times of intense movement. Extra-oral tissue’s purpose to keep the mouth adequately moist, would only prevent the evaporation of moisture.
Since this is the instance, the argument that open palates in theropods would lead to respiratory issues doesn’t hold up. Moreover, the constant need to move the jaws would cause extra stress on the supposed lower lip, leading to injury and infection due to the maxillary teeth constantly rubbing against them. A final point is that one may say that many modern animals may pant to vent excess heat but no terrestrial animal today has reached such sizes of the likes of torvosaurus. Animals of such size would need to evolve traits to help with cooling and extra-oral tissue are definitely not an advantage.
The theropod tooth cycle is another example on how lizard-like lips would pose as a grave disadvantage. Constant shedding of teeth (particularly the maxillary teeth) would lead them to accidentally lodge themselves in the ‘trough’ of the lower lips, given that if theropods were to have the same model of extra-oral tissue as squamates. Even though normally, teeth shed through feeding, it wouldn’t be an abnormality for this to occur. Squamate teeth are much smaller compared to the likes of theropod teeth and many lack the ‘trough’ on the lower lips. If theropod teeth did get stuck between the lower lip this could lead to infection, especially from the replacement tooth pushing down against it. This really isn’t a great evolutionary advantage.
Moreover, theropod tooth replacement mechanisms would push the tooth due to be replaced further down the dentary eventually passing the foramina (from which the lower lips are supposedly attached). This condition is seen clearly Tyrannosaurus rex Stan (Fig. 4) where the maxillary teeth extend past the dentary foramina. Stan’s right dentary also highlights the tooth replacement process particularly well, with several replacement teeth moving into position within existing teeth.

The phylogenetic argument doesn't mean what people seem to think it means, and even if it did, it doesn't describe the animals' evolutionary history according to the fossil record. Look at it this way: we can test the logic of "if it's an ancestral feature, it must show up on derived animals" against other points of the fossils known for both taxa. Look at how many unguals Yutyrannus and Dilong bear on their hands. Is that the same number of unguals found in tyrannosauridae? Look at the shape of their skulls--so allosaur- or carcharodontosaur-like that a few scientists have floated the idea Yutyrannus should belong in allosauroidea somewhere. Does that mean we should reject the skull shape of tyrannosaurids because it doesn't match our expectations?
Phylogenetics, when used properly, can produce some pretty robust hypotheses, but they're still hypotheses, and in order to use it right, you have to understand some fundamental conceptual limitations.
1. Phylogenetics CANNOT describe ancestry. This is why we always call related taxa "sisters," never "mother/daughter," even in cases where anagenesis has been proposed.
2. Species concept in paleontology is morphology-based, unlike other biological sciences. It therefore demands taking fossil phylogenies with a grain of salt, as it were.
3. Phylogenetic brackets require using the closest related animals possible for the strongest hypotheses. The closest brackets to any given theropod for are archosaurs and birds at this point.
4. Phylogenetic brackets have to actually bracket the taxon in question.
5. Most importantly, organisms produce variety. The equation of evolutionary theory does not work otherwise, rendering phylogeny moot.
It doesn’t matter that almost all animals have lips. That is a logical fallacy of hasty generalisation.
And phylogenetic argument here does not support the hypothesis of "lips" being present in archosauria as a whole, not only theropods.
Many people in support of lips claim the following:
‘Living dinosaurs (birds) and their closest relatives (crocodiles) do not serve as good comparisons, as they are both extremely specialised at what they do and show morphology that does not appear until late in their evolution’
So liplessness in crocodilians is a specialization? For what? How did they conclude this? How do they know they didn't inherit the lipless condition from their terrestrial ancestors (perhaps as far back as their most recent common ancestor with dinosaurs)? It's kind of a strange argument to make. It'd be like future paleontologists arguing that since old world chameleons are specialized lizards, lizards alive in their time can't be used to reconstruct their appearance. They're weird, so don't give them scales! There needs to be an argument for why liplessness in crocs is a specialization unique to their lifestyle, and therefore shouldn't be extended outside their clade.